Lichtwahrnehmung und Lichtnutzung
Inhalt
Übergeordneter Artikel:Dieser Text:
VB230.1
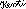 Moleküle die der Wahrnehmung und Nutzung von Licht dienen
Moleküle die der Wahrnehmung und Nutzung von Licht dienenVB230.1.1
Opsine, die Lichtrezeptoren aller Tiere, die näher mit uns verwandt sind als die SchwämmeVB230.1.1.1
Die OpsineVB230.1.1.2
Die Opsine der WirbeltiereVB230.1.1.3
Die Opsine der mehrzelligen Tiere und ihre Verwurzelung in den G-Protein gekoppelten RezeptorenVB230.1.1.3.1
Ein Stammbaum der OpsineVB230.1.1.3.2
DeuterostomiaVB230.1.1.3.2.1
VB230.1.1.3.2.2
Purpur-Seeigel (Strongylocentrotus purpuratus)VB230.1.1.3.2.3
Amphiura constrictaVB230.1.1.3.2.4
VB230.1.1.3.2.
VB230.1.1.3.3
ProtostomiaVB230.1.1.3.3.1
Der Unterschied zwischen Protostomia und DeuterostomiaVB230.1.1.3.3.2
Prostheceraeus vittatusVB230.1.1.3.3.3
Die Pazifische Auster (Crassostrea gigas) ist nicht völlig blindVB230.1.1.3.3.
VB230.1.1.3.4
Tiere, die keine Bilateria sindVB230.1.1.3.4.1
EinführungVB230.1.1.3.4.2
Trichoplax adhaerens - zwei Zellschichten, ein paar Fasern, keine Augen, aber zwei OpsineVB230.1.1.3.4.3
Die Seeanemone Nematostella vectensis hat Opsine aber keine AugenVB230.1.1.3.
VB230.1.2
Mikrobielle Rhodopsine: Rhodopsinähnliche Moleküle als lichtgetriebene IonenpumpenVB230.1.2.1
Stammbaum der Opsinähnlichen MoleküleVB230.1.2.2
VB230.1.3
Cryptochrome zur Farbwahrnehmung im blauen oder ultravioletten BereichVB230.1.3.1
Cryptochrom im Auge der Larve eines SchwammsVB230.1.4
ChlorophyllVB230.1.
vVB230.2
Typen von LichtsinneszellenVB230.3
Bauformen von AugenVB230.3.1
Augen gehen leichter verloren als sie neu entstehenVB230.3.2
Einzelne LichtsinneszellenVB230.3.2.1
Hautlichtsinn EinführungVB230.3.2.2
Lichtwahrnehmung in der Haut des MenschenVB230.3.2.3
Die gewöhnliche Krake (Oktopus vulgaris)VB230.3.2.4
VB230.3.3
FlachaugenVB230.3.4
PigmentbecheraugenVB230.3.5
GrubenaugenVB230.3.6
LochaugenVB230.3.7
LinsenaugenVB230.3.7.1
EinleitungVB230.3.7.2
Warnowiaceae - Einzeller mit LinsenaugenVB230.3.7.3
Terebratalia transversa - Linsenaugen aus nur zwei Zellen VB230.3.7.4
Stemmata - kleine Linsenaugen bei InsektenlarvenVB230.3.7.5
Das Rhopalium von Tripedalia cystophora als Beispiel für Linsenaugen bei QuallenVB230.3.7.6
SpringspinnenVB230.3.7.7
Der Gewöhnliche Tintenfisch (Sepia officinalis), die Lichtwahrnehmung der Chromatophoren und ein schnell wechselndes Aussehen für Tarnung, als Drohung und zur VerständigungVB230.3.7.8
Malacosteus niger - ein Tiefseefisch mit einer Laterne, deren Licht für andere Tiere der Tiefsee unsichtbar ist und den passenden Augen, um damit zu sehenVB230.3.8
SpiegelaugenVB230.3.9
Facettenaugen (Komplexaugen)VB230.
Quellen1. Moleküle die der Wahrnehmung und Nutzung von Licht dienen
1.1 Opsine, die Lichtrezeptoren aller Tiere, die näher mit uns verwandt sind als die Schwämme
1.1.1 Die Opsine
Die Opsine oder Rhodopsine, die Sehfarbstoffe, dienen der Zelle als Sensoren zur Lichtwahrnehmung. Rhodopsin ist nicht klar definiert und wird manchmal für alle Sehfarbstoffe verwendet, manchmal aber auch eingeschränkt auf die Farbstoffe für das Schwarzweißsehen. Ich benutze bei Wirbeltieren im folgenden das Wort Opsine für alle Sehfarbstoffe und verwende Rhodopsin eingeschränkt auf den Farbstoff für das schwarz-weiß-sehen in Dämmerung und Nacht der Wirbeltiere. Bei Tieren, die keine Wirbeltiere sind, gibt es diese Unterscheidung so nicht.Das erste Opsin, dessen genaue räumliche Struktur aufgeklärt wurde, war das Rhodopsin des Rindes33., daher wird das hier auch gezeigt. Andere Opsine sehen sehr ähnlich aus, so daß ich zumindest auf den ersten Blick keinen Unterschied erkenne, auch wenn sie sich in Details unterscheiden, da diverse Aminosäuren im Laufe der Evolution verändert wurden.

|
|
Bildquelle: 32. Rhodopsin des Rindes |
|
|
Bildquelle: 35.1
|
|
|
Bildquelle: 34.
|
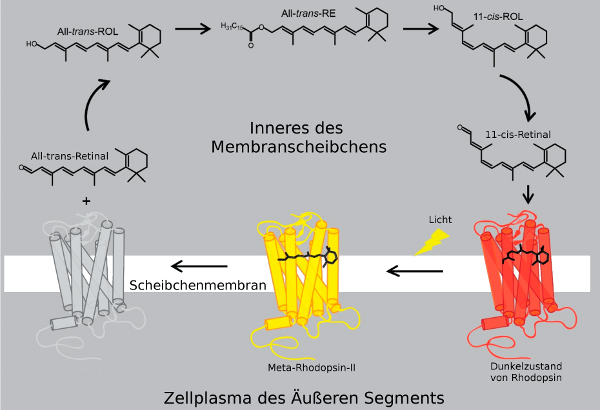
- Das eintreffende Photon (hv) wird absorbiert und aktiviert das Rhodopsin durch eine Konformationsänderung in der Scheibenmembran zu Meta-II-Rhodopsin (R*).
- Als nächstes hat Meta-II-Rhodopsin wiederholten Kontakt zu Transducin-Molekülen.
- Dies katalyisiert die Aktivierung zu G* durch die Freisetzung von gebundenem GDP und bindet dann freies GTP. Die Alpha- und Gamma Untereinheiten von G* binden an die hemmende Gamma Untereinheit der Phosphodiesterase (PDE), was seine Alpha- und Beta Untereinheiten aktiviert.
- Aktiviertes PDE hydrolysiert cGMP zu GMP. Reduzierte Mengen von freiem cGMP führen dazu, dass die Ionenkanäle in der Zellmembran geschlossen werden und weiteren Einstrom von Na+ und Ca2+ verhindern. Die Photorezeptorzelle hyperpolarisiert.
- Erst wenn die Phosphodiesterase (PDE) nicht mehr ständig neu aktiviert wird, weil das Opsin wieder in den Dunkelzustand zurückgekehrt ist, kann die Guanylyl-Cyclase (GC) die Oberhand gewinnen und genug cGMP prouzieren, welches als second messenger, als zweiter Botenstoff fungiert und den Dunkelzustand wiederherstellt, so daß die Ionenkanäle wieder öffnen.
O7.19.2.2.3.2
Die Lichtaktivierungskette
1.1.2 Die Opsine der Wirbeltiere
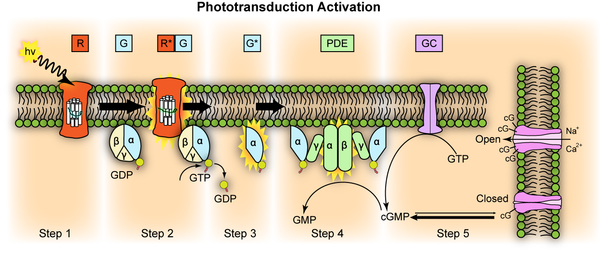
Bei den Wirbeltieren gibt es 5 Gruppen von zum Sehen verwendeten Opsinen. 4 davon dienen üblicherweise dem Farbensehen in den Zäpchen, einer dem sehen in Dämmerung und Dunkelheit in den Stäbchen. In der folgenden Graphik steht lw (für langwellig - rot markiert) für ein Opsin das im langwelligen Bereich am Besten absorbiert. Die meisten untersuchten Arten sehen damit im roten Bereich, einige im grünen Bereich. Bei den Menschenaffen hat sich das Gen verdoppelt und eine Variante ist mutiert, so daß wir damit grün und rot sehen können, bei dem Zebrabärbling ist das ebenfalls geschehen und er unterscheidet hiermit zwei verschiedene Rottöne. Das zweite Opsin dient bei den meisten Wirbeltieren zur Wahrnehmung von UV-Licht und heißt deshalb UVS oder aus sw1 (für short wave 1) einige Tiere sehen damit violett, der Mensch sieht damit blau. Ein weiteres Opsin dient bei den Wirbeltieren, die es besitzen zur Wahrnehmung von blauem Licht. Bei den Säugetieren und damit auch beim Menschen ist es verlorengegangen. RH2 ist so abgekürzt, weil es dem Rhodopsin auffallend ähnlich ist und dient bei den Arten, die es besitzen der Wahrnehmung von grünem Licht. RH (Rhodopsin) dient dem Sehen bei Dämmerung und Dunkelheit. 12.Warum wir so viele sehr ähnliche miteinander verwandte Opsine haben, wird verständlich, wenn man weiß, daß sich bei der Entstehung der Wirbeltiere das gesamte Genom zwei mal verdoppelt hatte. Frühe Wirbeltiere hatten also alle Gene ihrer Nicht-Wirbeltier-Vorfahren vier mal. Danach haben sich die vier Exemplare eines Wirbeltiergens jeweils auseinander entwickelt und sind teilweise auch verloren gegangen, weil sie nicht vier mal benötigit wurden. Einer dieser vier ursprünglichen Opsine hat sich dann noch einmal verdoppelt (RH/RH2 im Bild unten) und aus einer der Versionen entstand das Rhodopsin, was wir zum Dämmerungssehen verwenden.70. Diese fünf ursprünglichen Opsine und die Aufteilung in Stäbchen zum sehen bei heller Beleuchtung und Zäpfchen zum sehen bei schlechter Beleuchtung, ist so schon bei den Neunaugen (Petromyzontiformes) vorhanden, die sich abgespalten haben, bevor wir unseren Kiefer zum Kauen entwickelt haben.71. Allerdings erscheinen die Photoreezeptorzellen mit der Funktion der Zäpchen eher länglich wie Stäbchen bei uns und die die als Zäpfchen für das sehen bei Dunkelheit fungieren eher kurzs und dick wie Stäbchen72..

|
|
Bildquelle: 12.1
Stammbaum der Opsine der Wirbeltiere |
Die Ahnen der Säugetiere, die Therapsiden, waren Tetrachromaten. Sie hatten vier Sorten Zäpfchen mit jeweils unterschiedlichen Sehfarbstoffen, wie es heute beispielsweise die meisten Vögel und Reptilien auch haben. Sie hatten Zäpfchen für grün (RH2), violett (UVS/sw1), rot (lw) und blau (sw2). Erst den Säugetieren ist eine dieser Zäpfchensorten verlorengegangen, nämlich die für grün (RH2), daher ist oben kein Säugetier im grünen Kästchen zu finden. Das Opsin für violett (UVS/sw1) hat sein Wahrnehmungsspektrum in den ultravioletten Bereich verschoben. Wie wir Menschen war der Ahne aller Säugetiere Trichromat, im Gegensatz zu uns konnte er jedoch ultraviolettes Licht sehen. Alle jetzt noch existierenden Säugetiere haben einen weiteren Farbstoff verloren. Bei den Monotrematen, zu denen Schuppen- und Schnabeltiere gehören, ist es der für Ultraviolett (UVS/sw1), sie können also nur noch blau und rot sehen. Bei den Eutheria, zu denen die Beuteltiere (Marsipialia) und die Plazentatiere (Plazentalia, Mensch und Maus gehören beide in diese Gruppe) gehören, ist der für Blau (sw2) verloren gegangen. Unsere gemeinsamen Ahnen konnten daher nur UV-Licht (UVS sw2) und Rot (lw) sehen, waren also ähnlich ausgestattet, wie rot-grün-farbenblinde Menschen, nur daß sie bis in den ultravioletten Bereich hinein sehen konnten. So sieht die Maus heute noch. Bei den höheren Primaten hat sich der ursprünglich für rot vorgesehene Farbstoff genetisch verdoppelt und etwas verändert so daß wir jetzt wieder Trichromaten sind. Das Zäpfchen für grün der Maus liegt in dem Bereich mit den Opsinen, die ursprünglich rot gesehen haben (lw), der Mensch hat hier sein Zäpfchen für rot und das für grün. Die Maus hat ein UV-Zäpfchen, beim Menschen ist aus dem Farbstoff für dieses Zäpfchen einer entstanden, der sein Wahrnehmungsmaximum im blauen Bereich hat, während das Opsin sw2, mit dem Monotrematen und unsere Ahnen blau sehen konnten, uns verloren gegangen ist.12., 14..
|
|
Bildquelle: 14.1 Evolution der Lichtsinneszellen der Säugetiere. |
 Zuzana Musilova et Al. haben 100 Knochenfische untersucht und stellten fast, daß sie meist noch mehr Opsine zum sehen verwenden als ihre Verwandten vom Land, nämlich im Schnitt 7. Dabei führt schwache Beleuchtung nicht zu weniger Farbtüchtigkeit, wohl aber zu einer Verschiebung des sichtbaren Spektrums weg von rot und gelb und hin zu grün und blau, die in der Tiefsee am stärksten vertreten sind. Elf der untersuchten Fische hatten mehrere Gene für Rhodopsine (RH), die in Stäbchen zum sehen bei schwachen Licht verwendet werden und es ist daher anzunehmen, daß sie in der sehr dunklen Tiefsee Farben sehen können13..
Zuzana Musilova et Al. haben 100 Knochenfische untersucht und stellten fast, daß sie meist noch mehr Opsine zum sehen verwenden als ihre Verwandten vom Land, nämlich im Schnitt 7. Dabei führt schwache Beleuchtung nicht zu weniger Farbtüchtigkeit, wohl aber zu einer Verschiebung des sichtbaren Spektrums weg von rot und gelb und hin zu grün und blau, die in der Tiefsee am stärksten vertreten sind. Elf der untersuchten Fische hatten mehrere Gene für Rhodopsine (RH), die in Stäbchen zum sehen bei schwachen Licht verwendet werden und es ist daher anzunehmen, daß sie in der sehr dunklen Tiefsee Farben sehen können13..

|
|
Bildquelle: 31. Diretmus argenteus fiel in einer Untersuchung zu den Opsinen der Tiefseefischen dadurch auf, daß er 38 Rhodopsine für Stäbchen und 2 Opsine für Zäpchen hatte, die durchweg am stärksten im blauen und grünen Bereich absorbieren diesen Bereich aber recht gleichmäßig abdecken.13. |
1.1.3 Die Opsine der mehrzelligen Tiere und ihre Verwurzelung in den G-Protein gekoppelten Rezeptoren
1.1.3.1 Ein Stammbaum der Opsine
Um herauszufinden, wie eng oder weit die verschiedenen Opsine miteinander verwandt sind haben Clemens Christoph Döring die Gensequenzen der Opsine möglichst vieler unterschiedlicher Tiere aus öffentlich zugänglichen Genbibliotheken herausgesucht und die Basenabfolge, die die Buchstaben des genetischen Alphabets bilden, miteinander verglichen. Wie man bei früheren ähnlichen Untersuchungen zu ähnlichen Themen schon herausgefunden hat, entstehen dabei Stammbäume der Opsingene, die teilweise die Verwandtschaftsverhältnisse der untersuchten Tiere zueinander widerspiegeln, teilweise aber auch vom Zeitpunkt einer Genduplikation an, zeigen wie sich die verdoppelten Opsine danach auseinanderentwickeln.
6.Diese Opsine gehören zu der Gruppe der G-Protein-gekoppelten-Rezeptoren, die hier verwendet wurden, um zu zeigen, wo die Wurzel des Stammbaums liegt.
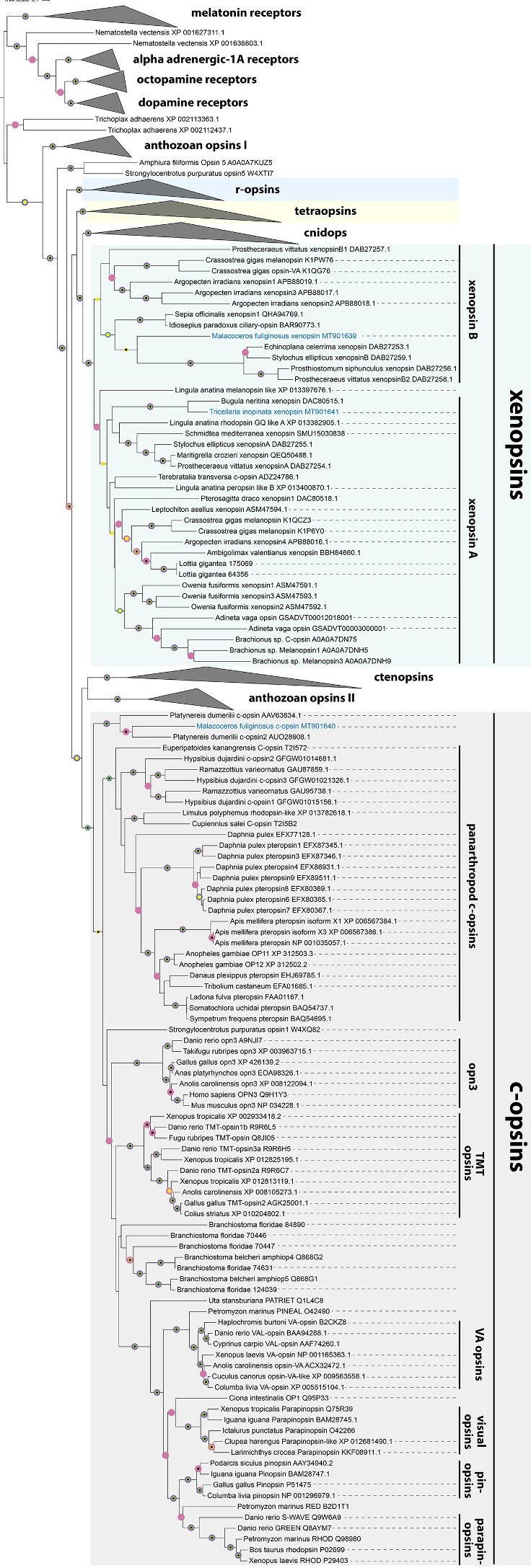
|
|
Bildquelle: 6.1
|
1.1.3.2 Deuterostomia
1.1.3.2.1
1.1.3.2.2 Purpur-Seeigel (Strongylocentrotus purpuratus)

|
|
Bildquelle: 83. Purpur-Seeigel (Strongylocentrotus purpuratus) ist ein Seestern (Echinoidea) |
1.1.3.2.3 Amphiura constricta

|
|
Bildquelle: 10. Amphiura constricta gehört zu den Schlangensternen (Ophiuroidea) |
1.1.3.2.
1.1.3.3 Protostomia
1.1.3.3.1 Der Unterschied zwischen Protostomia und Deuterostomia
Der Urahn der Bilateria, der existierte bevor die Deuteriostomia sich von den Protostomia abgetrennt hatte besaß mindestens neun Opsine. 85.1.1.3.3.2 Prostheceraeus vittatus

|
|
Bildquelle: 7. Prostheceraeus vittatus ist wasserlebend und sieht ein wenig wie eine Nacktschnecke aus, gehört aber zu den Strudelwürmern (Turbellaria) unter den Plattwürmern (Plathelminthes) |
1.1.3.3.3 Die Pazifische Auster (Crassostrea gigas) ist nicht völlig blind
Ich habe Muscheln immer für blinde Tiere gehalten, weil sie keine für mich erkennbaren Augen hatten. Tatsächlich haben aber auch diejenigen Muscheln Opsine, die keine erkennbaren Augen haben, wie das Beispiel der Pazifischen Auster (Crassostrea gigas) zeigt.

|
|
Bildquelle: 8. Crassostrea gigas ist eine Auster (Ostreidae) und zählt damit zu den Muscheln (Bivalvia) |
1.1.3.2 Tiere, die keine Bilateria sind
1.1.3.2.1 Der Unterschied zwischen Bilateria und den älteren Tiergruppen
Die Bilateria oder Zweiseitentierchen werden so genannt, weil ihre rechte Seite ein Spiegelbild ihrer linken Seite ist. Natürlich gibt es spätere Abwandlungen dieses ursprünglich symmetrischen Bauplans, die nicht mehr ganz so symmetrisch sind. Während wir beispielsweise äußerlich weitgehend symmetrisch aussehen, haben wir nur eine Leber und auf der gegenüberliegenden Seite die Milz statt dort eine zweite Leber zu besitzen. Tiere, die nicht zu den Bilateria zählen, sind nicht zweiseitig symmetrisch, wie wir das sind, sondern besitzen einen meist unregelmäßigeren Bauplan. Für unsere Begriffe sehen sie oft gar nicht so wirklich wie Tiere aus, sondern erinnern uns beispielsweise an Blumen oder ... Putzlappen. Zumindest erinnert mich Trichoplax adhaerens am ehesten an einen Lappen. Trotz ihres für unsere Begriffe wenig tierähnlichen Bauplans haben sie mit uns normalerweise durchaus Opsine gemein. Es ist also anzunehmen, daß sie in irgendeiner Form Licht wahrnehmen.
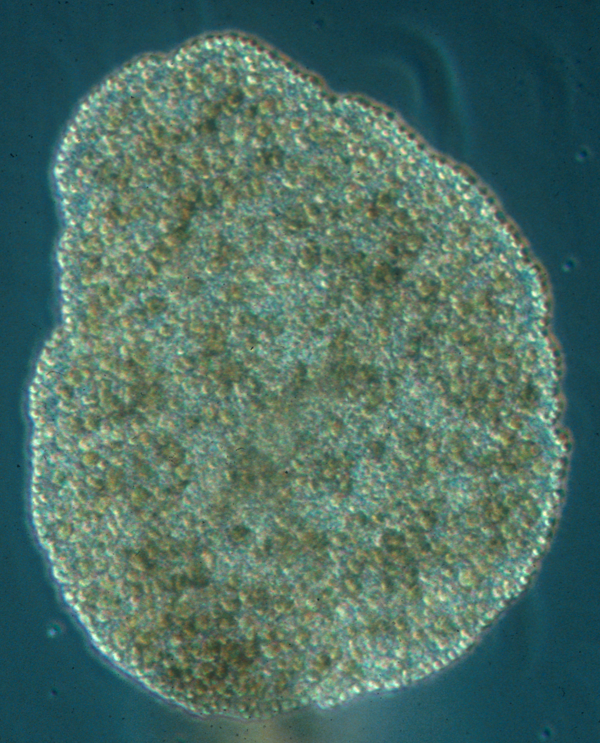
1.1.3.2.2 Trichoplax adhaerens - zwei Zellschichten, ein paar Fasern, keine Augen, aber zwei Opsine
|
|
Bildquelle: 11.1 Trichoplax adhaerens |
Trichoplax hat man wegen seines einfachen Bauplanes für ein sehr primitives Tier gehalten, doch es ist tatsächlich näher mit uns verwandt, als die Schwämme18., die mit uns ein Entwicklunsstadium gemein haben, das unserem Blastulastadium entspricht und offensichtlich nicht nur eine oberflächliche Ähnlichkeit hat, sondern tatsächlich auf denselben chemischen Mechanismen beruht und daher homolog ist23.. Das heißt, daß die Placozoa von Vorfahren abstammen, die komplexer aufgebaut sind sind als diese Art und erst nachträglich wieder diese einfache Bauform angenommen haben, die wir heute bei ihnen vorfinden.
1.1.3.2.3 Die Seeanemone Nematostella vectensis hat Opsine aber keine Augen

|
|
Bildquelle: 9. Nematostella vectensis |
Nematostella vectensis besitzt keine sichtbaren Augen, hat aber trotzdem über zwanzig verschiedene Opsine, von denen zwei zu der Clade "Cnidops" zählen, während der Rest unter "anthozoan opsins II" zu finden ist.26.
1.2 Mikrobielle Rhodopsine: Rhodopsinähnliche Moleküle als lichtgetriebene Ionenpumpen
1.2.1 Stammbaum der Opsinähnlichen Moleküle
Bei den Stammbäumen zu den Opsinen der Tiere hat man die Basenabfolge der zugehörigen Gene angesehen, sie mit Hilfe von Computerbrogrammen miteinander verglichen und daraus geschlossen, wie nahe die verschiedenen Varianten des Opsins miteinander verwandt sind. Wenn man zu immer kleineren Lebewesen und immer weiter im Stammbaum zurückgeht, funktioniert dieser Trick nicht mehr. Ab irgend einem Punkt haben sich die Basenabfolgen da, wo man sie ungestraft ändern kann, so oft hin- und zurückverändert, daß man nicht mehr sagen kann, was denn nun am engsten mit was verwandt ist. Nur die für die Funktion des Opsins wichtigen Teile der jeweiligen Gene sind erhalten geblieben.

|
|
Bildquelle: 17.1
|
1.3 Cryptochrome zur Farbwahrnehmung im blauen oder ultravioletten Bereich
Cryptochrome gehören zu einer Molekül-Familie, die im blauen oder ultravioletten Bereich zur Farbwahrnehmung fähig ist. Außerdem zählen dazu Photolyasen, die Lichtenergie benutzen, um DNA-Schäden zu reparieren. Cryptochrome wurden in den Augen von Amphibien, Insekten und Säugetieren festgestellt, sind auch in Weichtieren verbreitet und spielen eine Rolle bei der Kontrolle des Tag-Nacht-Rhythmusses und Kompass-Navigation bei Vögeln und Insekten.49., 86.Cryptochrom im Auge der Larve eines Schwamms

|
|
Bildquelle: 23.2 Ausgewachsener Schwamm von Amphimedon queenslandica. |
VB218.6.6.4
Mikrotubuli in Zilien und FlagellenBeim Seßhaft werden, falten sich der Jungschwamm ähnlich wie der menschliche Embryo ein, so daß ein Hohlraum entsteht, der an das menschliche Blastula-Stadium erinnert. Diese Entwicklung wird auch durch dieselben Signalgene gesteuert, die diese Funktion auch in der menschlichen Embryonalentwicklung hat23.. Das Cryptochrom Aq-Cry2 dient bei der Parenchymella des Schwamms Amphimedon queenslandica der Lichtwahrnehmung.22.
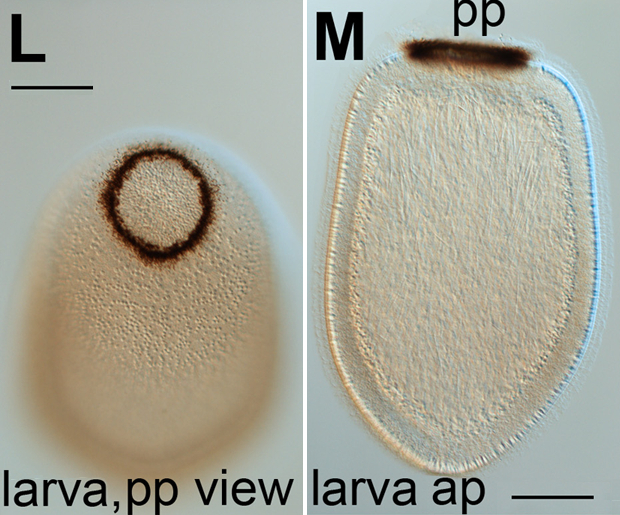
|
|
Bildquelle: 23.1 Parenchymella von Amphimedon queenslandica. Der dunkle Pigmentring dieser Larve fungiert als Auge, verwendet aber kein Opsin sondern das Cryptochrom Aq-Cry2 zur Lichtwahrnehmung. |
1.4 Chlorophyll
2. Typen von Lichtsinneszellen
Bakterien und Archaeen mit Lichtwahrnehmung
Einzelne eukaryotische Zellen mit Lichtwahrnehmung
Chlamydomonas reinhardtii
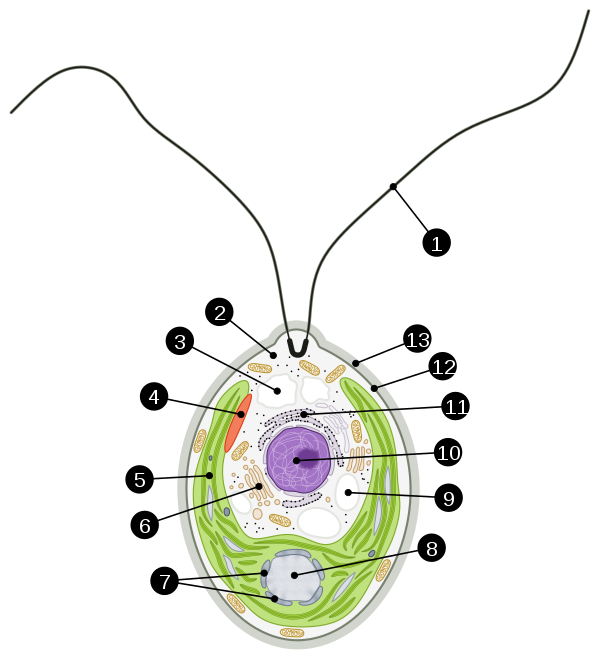
|
|
Bildquelle: 16.
Chlamydomonas reinhardtii |
VB218.6.6.4.4
Die Flagellen der EukaryotenDie Alge besitzt sieben verschiedene Rhodopsine von dem Typ wie sie Bakterien besitzen und die Chlamyopsine genannnt werden.
Sehende Spermien
Einfache Mehrzeller
Volvox
Lichtsinneszellen in Augen von Tieren
Stäbchen und Zäpfchen im Auge der Wirbeltiere
Wie Opsine den Seheindruck chemisch im Zusammenhang mit anderen Molekülen erzeugen, ist in folgendem Abschnitt am Beispiel der Lichtaktivierungskette, wie sie beim Menschen funktioniert, beschrieben.O7.19.2.2.3.2
Die Lichtaktivierungskette
Lichtsinneszellen im dritten Auge von Wirbeltieren
Lichtsinneszellen in den Facettenaugen der Insekten
Lichtsinneszellen in den Ozellen der Insekten
Lichtsinneszellen bei Pflanzen
3. Bauformen von Augen
3.1 Augen gehen leichter verloren als sie neu entstehen
Augen gehen leichter verloren als sie entstehen. In Muscheln sind aus einzelnen lichtempfindlichen Zellen am Mantelrand mindestens fünf mal unabhängig voneinander kompolexere Augentypen entstanden. Bei den Pterioidea entstanden Augen die wie eine Kappe auf einer pigmentierten Mantelfalte sitzen und dadurch eine ungefähre Einschätzung erlauben, woher das Licht kommt. Bei den Pectinoidea entstanden Spiegelaugen. Bei den Limida entstanden Linsenaugen. Bei den Arcida entstanden Pigmentbecheraugen, die bei einigen ihrer Untergruppen zu Komplexaugen gruppiert wurden. All diese Augentypen gingen sehr häufig wieder verloren. 88.
3.2 Einzelne Lichtsinneszellen
3.2.1 Hautlichtsinn Einführung
Die einfachsten „Augen“ sind lichtempfindliche Sinneszellen auf der Außenhaut, die als passive optische Systeme funktionieren. Sie können nur erkennen, ob die Umgebung hell oder dunkel ist. Man spricht hier von Hautlichtsinn.19., 20.
3.2.2 Lichtwahrnehmung in der Haut des Menschen
Wie sich ein Haut-Licht-Sinn anfühlt, wissen wir durchaus, denn denn in der menschlichen Haut werden alle Opsine von Stäbchen (OPN2) und Zäpfchen (OPN1) mit denen wir im auge sehen, sowie OPN3 (Encephalopsin oder Panopsin) und OPN5 (Neuropsin) produziert29., 30.. In der Haut wird auch Melanopsin (OPN4) exprimiert und hat dabei in den Melanozyten die Funktion die Hautbräunung zu steuern40.. Melanozyten enthalten also OPN1,2,3,4,540., 43. und sind auch mit dem Nervensystem verbunden und können ihre Lichtwahrnehmung daher auch daran weiterleiten42.. Ich zumindest kann mit geschlossenen Augen durchaus fühlen, von woher die Sonne scheint und wo ein heißer Ofen steht und das geht mit der nackten Haut deutlich genauer als wenn ich angezogen bin.Daneben hat Melanopsin in der Haut noch an anderer Stelle Funktionen: Im Unterhautfettgewebe ist Melanopsin dafür verantwortlich, daß wir im Winter, wenn die Haut weniger Licht erhält, ein dickeres Unterhautfettgewebe haben als im Sommer39.. Darüber hinaus wird Melanopsin in Blutgefäßen exprimiert, wo Beleuchtung dazu führt, daß die Adern sich ausdehnen41..
3.2.3 Die gewöhnliche Krake (Oktopus vulgaris)
Die gewöhnliche Krake gehört zu den Kopffüßlern (Cephalopoda) und diese wie Schnecken und Muscheln zu den Weichtieren (Mollusca). Er hat eine viel einfacher aufgebaute Retina. Die Sinneszellen bestehen aus zwei Segmenten. Dem Glaskörper zugewendet ist der Teil mit den Rhabdomeren, 52.3.3 Flachaugen
Diverse Quallen und Seesterne besitzen flache Augen mit vielen nebeneinander liegende Lichtsinneszellen, die innen an eine Schicht aus Pigmentzellen anschließen können. Die Konzentrierung der Sinneszellen verbessert die Hell-Dunkel-Wahrnehmung.19., 20.3.4 Pigmentbecheraugen Die Sehzellen liegen vom Licht abgewandt (inverse Lage) in einem Becher aus lichtundurchlässigen Pigmentzellen. Das Licht kann nur durch die Öffnung des Bechers eindringen, um die Sehzellen zu stimulieren. Da daher immer nur ein kleiner Teil der Sehzellen gereizt wird, kann neben der Helligkeit auch die Einfallsrichtung des Lichts bestimmt werden. Solche Augen besitzen unter anderem Strudelwürmer und Schnecken.19., 20.
3.5 Grubenaugen
Das Grubenauge unterscheidet sich vom Pigmentbecherauge durch die dem Licht zugewandte (everse) Lage der Sinneszellen und dadurch, dass die Grube mit Sekret gefüllt ist. In der Grube bilden die Sehzellen eine Zellschicht, die innen an eine Schicht von Pigmentzellen anschließt. Es ist also eine Weiterentwicklung des Flachauges. Es ermöglicht auch die Bestimmung der Intensität und der Einfallsrichtung des Lichts.19., 20.3.6 Lochaugen
Lochaugen sind verbesserte Grubenaugen. Die Öffnung der Grube ist nur noch ein kleines Loch und der Hohlraum ist vollständig mit Sekret gefüllt. Durch die erhöhte Anzahl der Sehzellen ist nun auch Bildsehen möglich. Das Bild ist jedoch lichtschwach und nur schemenhaft. Diesen Typ findet man bei niederen Tintenfischen. Eine Abart des Lochauges ist das Blasenauge, bei dem die Öffnung von einer durchsichtigen Haut bedeckt ist (Vorkommen bei machen Schneckenarten). Das Sekret kann auch zu einer einfachen Art von Linse verfestigt sein (bei Weinbergschnecken der Fall). Diese Modifikationen verbessern das Bild geringfügig.19., 20.3.7 Linsenaugen
3.7.1 Einleitung
Im Tierreich gibt es viele verschiedene Formen der Linsenaugen. die auswertung der Bilder beginnt oft schon im Auge.3.7.2 Warnowiaceae - Einzeller mit Linsenaugen
Bei der Frage, was die kleinsten Linsenaugen sind, würden sicherlich die meisten Menschen genau wie ich falsch raten und vermuten, daß sie vielleicht bei Schnecken oder ähnlichen mehrzelligen Tieren zu finden sein könnten. Tatsächlich wurden die kleinsten Linsenaugen bei Einzellern gefunden, die zu den Warnowiaceae gehören.
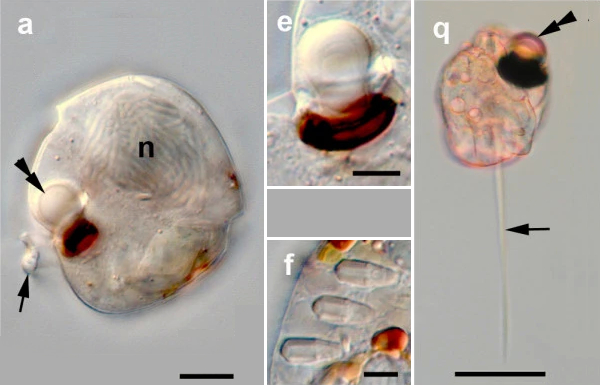
|
|
Bildquelle: 2.1 Augen bei den Warnowiaceae: a-f gehören zur Gattung Proterythropsis, e ist ein Ozelloid, bei f handelt es sich um eine Nesselkapsel oder Nematocyste, q gehört zur Gattung Erythropsidinium. Das schwarze n markiert den Zellkern, die Doppelpfeile zeigen auf Linsenaugen. In a repräsentiert der schwarze Strich eine Länge von 10 μm, in e, f und q 5 μm |
VB216.5.3.3
Endosymbiosebeziehung von Blaualgen (Cyanobacteria) als Blattgrünkörperchen (Chloroplasten) in Pflanzen (Plantae) und SchneckenDas Ozelloid ist in der Lage sich an unterschiedliche Lichtverhältnisse anzupassen, indem das Melanosom sich zusammenzieht oder streckt.1., 2.

|
|
Bildquelle: 1.1 Durchlichtelektronenmikroskopische Aufnahmen des Ocelloids, die zeigen, wie es sich bei veränderter Helligkeit verformt
|
3.7.3 Terebratalia transversa - Linsenaugen aus nur zwei Zellen

|
|
Bildquelle: 87. Erwachsenes lebendes Exemplar von Terebratalia transversa |
Nach einer Zelle kommen zwei - jedenfalls wenn man einfach zählt. Die Larve von Terebratalia transversa hat tatsächlich Linsenaugen, die aus nur zwei Zellen bestehen, von denen die eine eine Linse bildet, während die andere einen Pigmentbecher aus ein paar Pigmentkörnern enthält. Die Zilien beider Zellen nehmen in dem als Retina dienenendem Zwischenraum das Licht wahr, das dort hineinfällt. Diese Zilien sind auch das, was an dem Tierchen ungewöhnlich ist, denn es hat damit den Photorezeptortyp den wir von Wirbeltieren - beziehungsweise der übergeordneten Gruppe Deuterostomia - kennen und nicht den, den wir von Protostomia, zu denen Terebratalia transversa gehört, erwarten. 84.
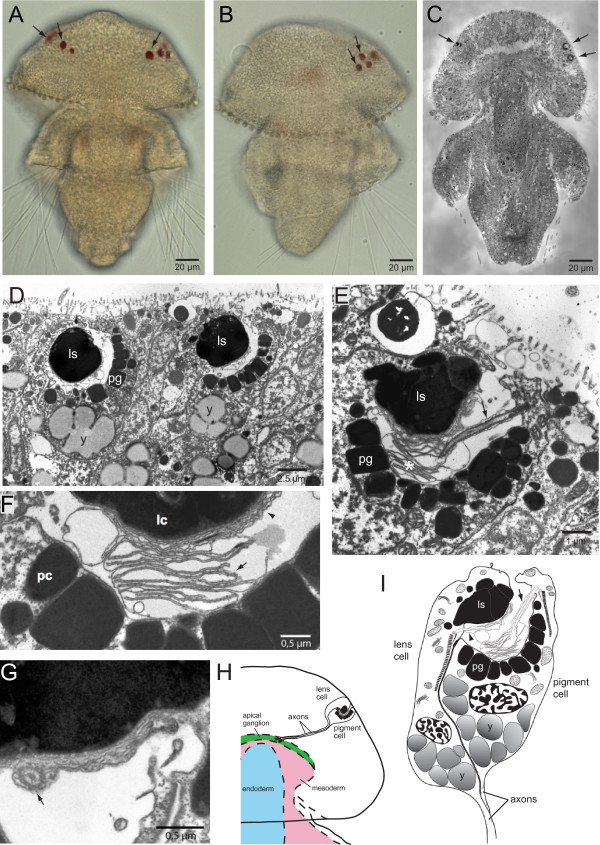
|
|
Bildquelle: 84.1
Die Larve von Terebratalia transversa.
|
3.7.4 Stemmata - kleine Linsenaugen bei Insektenlarven

|
|
Bildquelle: 52. Ein Weibchen des Gemeinen Furchenschwimmers (Acilius sulcatus) holt Luft an der Wasseroberfläche, indem es eine frische Luftblase unter die Flügeldecken nimmt |

|
|
Bildquelle: 54. Larve des Gemeinen Furchenschwimmers (Acilius sulcatus) |
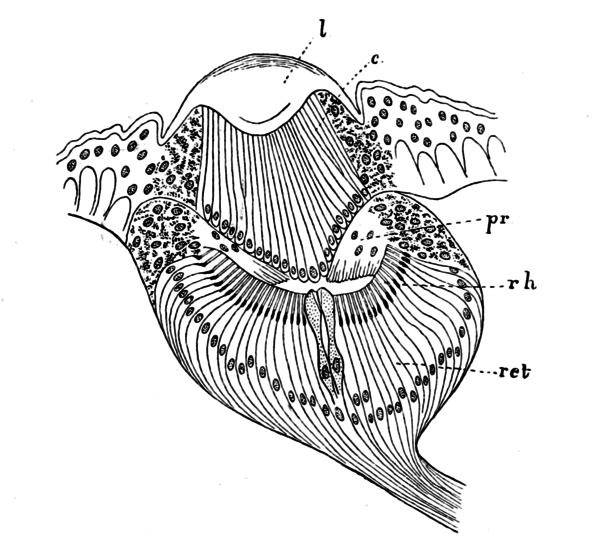
|
|
Bildquelle: 53. Das kleine Linsenauge oder Stemma der Larve der Gattung Acilius zu der der Gemeine Furchenschwimmer (Acilius sulcatus) gehört
|
3.7.5 Das Rhopalium von Tripedalia cystophora als Beispiel für Linsenaugen bei Quallen
Auch einige Quallen haben Linsenaugen, die sich gleich zu mehreren in den sogenannten Rhopalien befinden, kleine aus Tentakeln entstandene Anhänge, die häufig mehrere Augen und zusätzlich Statozysten zur Schwerkraftwahrnehmung enthalten.
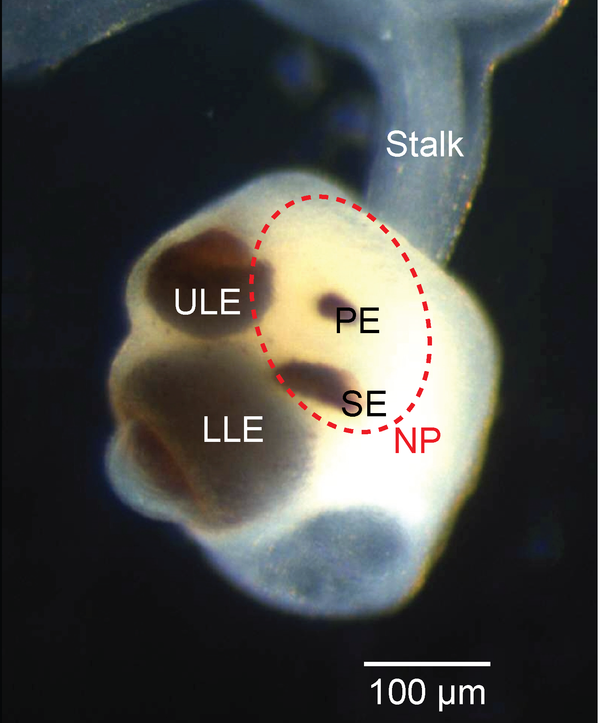
|
|
Bildquelle: 3.1 Das Rhopalium von Tripedalia cystophora ist nicht einmal einen Millimeter groß, enthält aber gleich mehrere Augen.
|
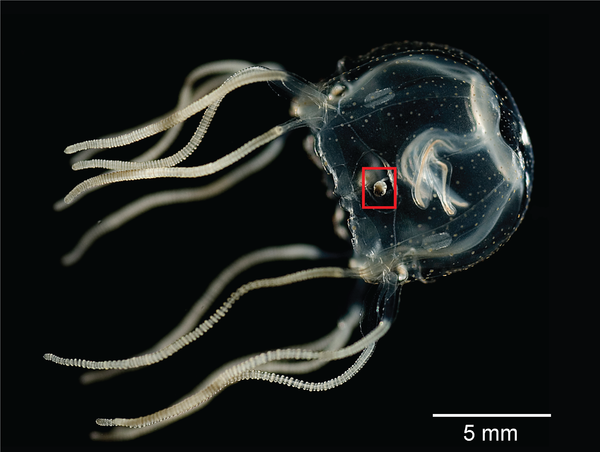
|
|
Bildquelle: 3.2 Tripedalia cystophora zählt zu den Würfelquallen (Cubozoa), um das Rhopalium ist ein rotes Kästchen gezeichnet. |
3.7.6 Springspinnen
Gleich von vier Augen angestarrt fühlt man sich, wenn man von einer Springspinne fixiert wird. Und falls man eine Fliege ist, sollte man vorsichtig sein. Sie hat durchaus recht gute Linsenaugen und kann auf 20cm Entfernung gezielt springen.

|
|
Bildquelle: 4. Zebraspringspinne (Salticus scenicus) - rechts etwas über den vierten äußeren Auge ist ein fünftes Auge zu sehen, das zur Seite und nach hinten sieht |
|
|
Bildquelle: 5.
|
3.7.7 Der Gewöhnliche Tintenfisch (Sepia officinalis), die Lichtwahrnehmung der Chromatophoren und ein schnell wechselndes Aussehen für Tarnung, als Drohung und zur Verständigung

|
| Bildquelle: 55. |
|
|
Bildquelle: 56. Auf Illustrationen sind die Fangarme des Gewöhnlichen Tintenfisches (Sepia officinalis) häufig zu sehen. |
|
|
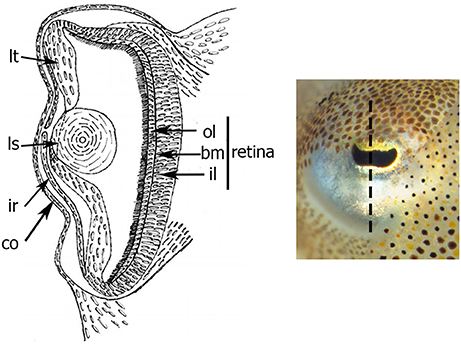
Bildquelle: 67. Auge des Gewöhnlichen Tintenfisches (Sepia officinalis). Die schwarz gepünktelte Linie zeigt wo das auge geschnitten wurde.
|
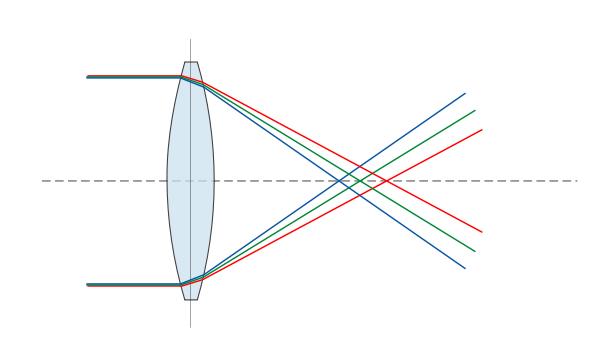
|
|
Bildquelle: 81. Eine Linse bricht - wenn man keine Gegenmaßnahmen ergreift - Licht unterschiedlicher Wellenlängen unterschiedlich stark. |

|
|
Bildquelle: 57. Gewöhnlicher Tintenfisch (Sepia officinalis), Ushaka Sea world, 10. Juli 2014, 15:07:27 |

|
|
Bildquelle: 58. Gewöhnlicher Tintenfisch (Sepia officinalis), Ushaka Sea world, 10. Juli 2014, 15:07:53 |

|
|
Bildquelle: 59. Gewöhnlicher Tintenfisch (Sepia officinalis) |

|
|
Bildquelle: 60. Gewöhnlicher Tintenfisch (Sepia officinalis) |

|
|
Bildquelle: 61.
|

|
|
Bildquelle: 63. Daß Die Anpassung an den Hintergrund nicht perfekt ist, merkt man bei künstliche Hintergründen. Tintenfische sind farbenblind, daher fühlt sich das gelb gepunktete 2 Monate alte Jungtier auf dem blauen Hintergrund gut angepaßt. Erkennbar ist hier auch eine sehr zottelige Hautstuktur. Ein grobes Schachbrettmuster führt auch beim Tintenfisch zu einem gröberen Muster dessen genaue Form durch die Körperformen vorgegeben ist, daher ergibt sich kein Schachbrett. In Bild C wurde das Tier erschreckt und zeigt eine Warnzeichnung, in der vorne die Augen durch dunkle Ränder betont werden und hinten zwei Augenflecken zu sehen sind. In D ist eine Mischung zwischen dem gepunkteten und dem gröberen Tarnmuster gezeigt. |
3.7.8 Malacosteus niger - ein Tiefseefisch mit einer Laterne, deren Licht für andere Tiere der Tiefsee unsichtbar ist und den passenden Augen, um damit zu sehen
Tiefseefische haben diverse Anpassungen an das sehen bei schwacher Beleuchtung, die nicht durch die Sonne sondern durch Biolumineszenz entsteht, wie große Augen, weite Pupillen, die teilweise noch größer sind als die Linse des Auges oder ein Tapetum lucidum hinter der Netzhaut, das das Licht zurück in die Netzhaut reflektiert, damit die Lichtsinneszellen mehr von dem Licht auffangen können. Die meisten Tiefseefische haben Retina die hauptsächlich Stäbchen enthält. Außerdem sind diese Stäbchen länger und haben daher mehr Membranscheibchen mit Rhodopsin und die Retina mancher Tiefseefische hat mehrere Lagen Stäbchen.36. Die meisten Tiefseefische sehen in Wellenlängen zwischen 470–490 nm am Besten weil in diesem Bereich der letzte Rest Tageslicht liegt, der noch bis in tiefere Lagen dringt und weil der größte Teil der Biolumineszens in denselben Wellenlängen liegt.

|
|
Bildquelle: 46. Beroe abyssicola gehört zu den tentakellosen Nuda unter den Rippenquallen (Ctenophora) und kann, wie viele andere Rippenquallen auch, durch Biolumineszens Licht erzeugen, das wie bei den meisten tiefseetieren im blaugrünen Bereich liegt. |

|
|
Bildquelle: 45. Malacosteus niger |
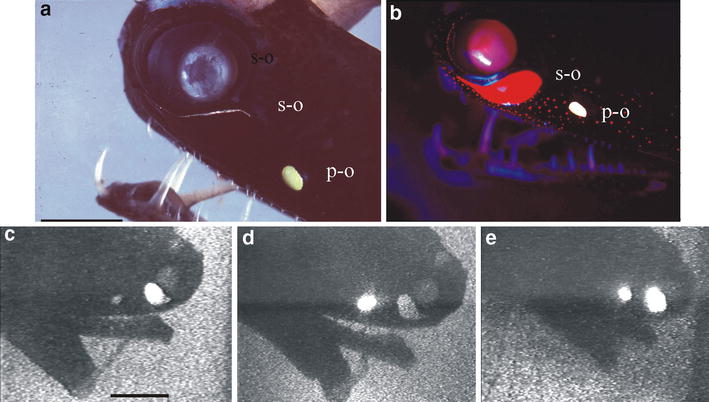
|
|
Bildquelle: 38.1
a) Kopf und Unterkiefer von Malacosteus niger im normalen Licht, s-o suborbitale Photophore, bei normaler Beleuchtung kaum erkennbar, p-o postorbitale Photophore
|
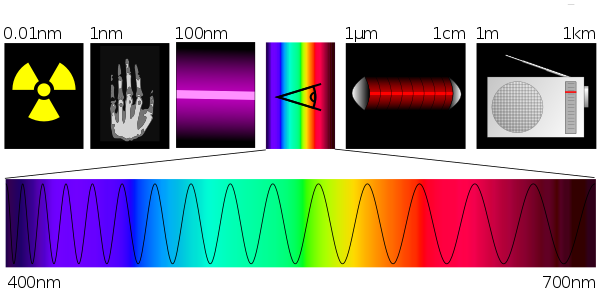
|
|
Bildquelle: 48. Wellenlängen elektromagnetischer Strahlung
|
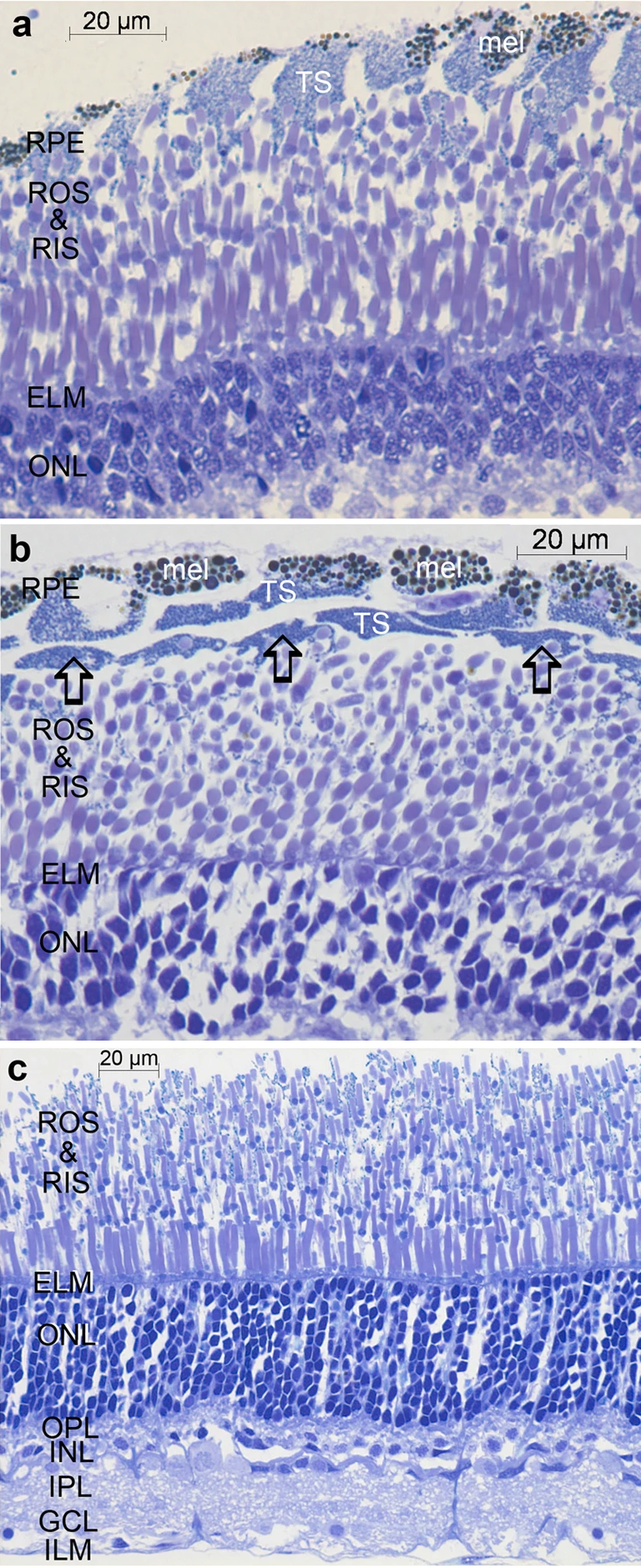
|
|
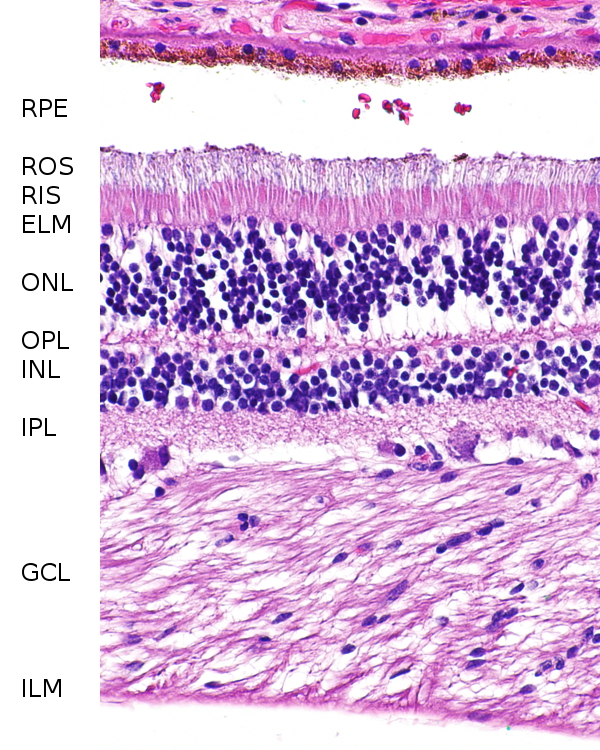
Bildquelle: 37.1 Retina von Malacosteus niger, gefärbt mit Toluidinblau.
a und b zeigt Ausschnitte der Retina vom Rand des Sichtfeldes, die vom Pigmentepithel (RPE) bis zu der äußeren Körnerschicht reichen, a ist radial geschnitten, der Schnitt zeigt also zum Zentrum der Retina und die Schichtung ist so nicht zu erkennen, b ist quer geschnitten und in dieser Darstellung scheinen die Pigmentepithelzellen mehrere übereinanderliegende Schichten zu bilden, von denen die äußerste das Melanin enthält.
|
|
|
Bildquelle: 44.
Zum Vergleich: Die zentrale Retina des Menschen nahe dem Austrittspunkt des Sehnervs. Bezeichnungen wie oben. Im Präparat ist das Pigmentepithel gerissen, die weiße Schicht gehört also nicht dahin, sondern die braun gepunkteten Pigmetzellen schließen direkt an die Äußeren Segmente (ROS) der Stäbchen an. |
O7.19.2.2
Das Sehen als Beispiel für eine Sinneswahrnehmung des Menschen3.8 Spiegelaugen
3.9 Facettenaugen (Komplexaugen)
Facettenaugen setzen sich aus einer Vielzahl von Einzelaugen (Ommatidien) zusammen. Jedes Einzelauge sieht nur einen winzigen Ausschnitt der Umgebung, das Gesamtbild ist ein Mosaik aus allen Einzelbildern. Die Anzahl der Einzelaugen kann zwischen einigen Hundert bis hin zu einigen Zehntausend liegen. Die Auflösung des Facettenauges ist durch die Anzahl der Einzelaugen begrenzt und ist daher weit geringer als die Auflösung des Linsenauges. Allerdings kann die zeitliche Auflösung bei Facettenaugen weit höher sein als bei Linsenaugen. Sie liegt etwa bei fliegenden Insekten bei 250 Bildern pro Sekunde, was etwa dem zehnfachen des menschlichen Auges entspricht. Dies verleiht ihnen eine extrem hohe Reaktionsgeschwindigkeit. Zu finden sind diese Augen bei Krebsen und Insekten.19., 20.Pecks Immenbremse (Xenos peckii) - Linsenaugen zu Komplexaugen zusammengesetzt

|
|
Bildquelle: 82.1 Rasterelektronenmikroskopisches Bild von den Augen der Männchen des Fächerflüglers Pecks Immenbremse (Xenos peckii) |

|
|
Bildquelle: 82.2 Schematische Darstellung der Augen der Männchen des Fächerflüglers Pecks Immenbremse (Xenos peckii) |
Quellen
-
Shiho Hayakawa,
Yasuharu Takaku,
Jung Shan Hwang, Takeo Horiguchi, Hiroshi Suga, Walter Gehring, Kazuho Ikeo, Takashi Gojobori: Function and Evolutionary Origin of Unicellular Camera-Type Eye Structure. In:
 PLoS One (2015) 10(3): e0118415.
PLoS One (2015) 10(3): e0118415.  PMID: 25734540, DOI: 10.1371/journal.pone.0118415 ( Volltext)
PMID: 25734540, DOI: 10.1371/journal.pone.0118415 ( Volltext)
- 1.1 ↑Bild VB23007.PNG:
Figure 2 (Änderungen zum Original: Kontraste erhöht, Buchstaben hervorgehoben) aus Shiho Hayakawa,
Yasuharu Takaku,
Jung Shan Hwang, Takeo Horiguchi, Hiroshi Suga, Walter Gehring, Kazuho Ikeo, Takashi Gojobori: Function and Evolutionary Origin of Unicellular Camera-Type Eye Structure. In: PLoS One (2015) 10(3): e0118415. PMID: 25734540, DOI: 10.1371/journal.pone.0118415 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 1.1 ↑Bild VB23007.PNG:
-
Mona Hoppenrath,
Tsvetan R. Bachvaroff, Sara M. Handy, Charles F. Delwiche, Brian S. Leander: Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences. In: BMC Evolutionary Biology, volume 9, Article number: 116 (2009) ( Volltext)
- 2.1
↑Bild VB230.JPG:
Verschiedene Aussschnitte aus Figure 1, in: Mona Hoppenrath, Tsvetan R. Bachvaroff, Sara M. Handy, Charles F. Delwiche, Brian S. Leander: Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences. In: BMC Evolutionary Biology, volume 9, Article number: 116 (2009) ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY 2.0 hochgeladen hast! Thank you very much!
- 2.1
↑Bild VB230.JPG:
Verschiedene Aussschnitte aus Figure 1, in:
-
Jan Bielecki,
Alexander K. Zaharoff, Nicole Y. Leung, Anders Garm, Todd H. Oakley: Ocular and Extraocular Expression of Opsins in the Rhopalium of Tripedalia cystophora (Cnidaria: Cubozoa). In: PLoS One 9(6): e98870 (2014). https://doi.org/10.1371/journal.pone.0098870 ( Volltext)
- 3.1 ↑Bild VB230.PNG:
File:Ropalio-tripedalia-cystophora-detail.png. Ausschnitt aus Figure 1. von Jan Bielecki, Alexander K. Zaharoff, Nicole Y. Leung, Anders Garm, Todd H. Oakley: Ocular and Extraocular Expression of Opsins in the Rhopalium of Tripedalia cystophora (Cnidaria: Cubozoa). In: PLoS One 9(6): e98870. https://doi.org/10.1371/journal.pone.0098870 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 3.2 ↑Bild VB23001.PNG:
File:Ropalio-tripedalia-cystophora-detail.png. Ausschnitt aus Figure 1. von Jan Bielecki, Alexander K. Zaharoff, Nicole Y. Leung, Anders Garm, Todd H. Oakley: Ocular and Extraocular Expression of Opsins in the Rhopalium of Tripedalia cystophora (Cnidaria: Cubozoa). In: PLoS One 9(6): e98870. https://doi.org/10.1371/journal.pone.0098870 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 3.1 ↑Bild VB230.PNG:
-
↑Bild VB23001.JPG:
File:Salticus scenicus, Nant Gwrtheyrn, North Wales, June 2015 3 (19379049041).jpg (oder hier) von Flickr-User Janet Graham
Vielen Dank, daß Du das Bild unter CC BY 2.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23002.PNG:
File:Sichtfeld Salticidae.jpg aus: David Edwin Hill: Use of location (relative direction and distance) information by jumping spiders (Araneae, Salticidae, Phidippus) during movement toward prey and other sighted objectives. In: Peckhamia 83.1, 28 October 2010, p. 14. ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY 3.0 hochgeladen hast! Thank you very much!
-
Clemens Christoph Döring,
Suman Kumar, Sharat Chandra Tumu, Ioannis Kourtesis, Harald Hausen: The visual pigment xenopsin is widespread in protostome eyes and impacts the view on eye evolution. In: eLife, 2020; 9: e55193. ( Volltext 1, 2)
- 6.1 ↑Bild VB23002.JPG:
Ausschnitt aus: Figure 2 "2C-opsins and xenopsins display type-specific conserved gene structures." aus Clemens Christoph Döring, Suman Kumar, Sharat Chandra Tumu, Ioannis Kourtesis, Harald Hausen: The visual pigment xenopsin is widespread in protostome eyes and impacts the view on eye evolution. In: eLife, 2020; 9: e55193. ( Volltext 1, 2)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 6.1 ↑Bild VB23002.JPG:
Ausschnitt aus:
-
↑Bild VB23003.JPG: Ausschnitt aus
Figure 4 "Prostheceraeus vittatus" aus: Carolina Noreña, Daniel Marquina, Jacinto Perez, Bruno Almon: First records of Cotylea (Polycladida, Platyhelminthes) for the Atlantic coast of the Iberian Peninsula. In: ZooKeys 404: 1-22 doi: 10.3897/zookeys.404.7122, Published: 22 Apr 2014 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
-
↑Bild VB23004.JPG:
File:Ostras 03-03.JPG von User:P.Lameiro von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23005.JPG:
File:Nematostella vectensis (I1419) 999 (30695685804).jpg und hier von Robert Aguilar, Smithsonian Environmental Research Center.
Vielen Dank, daß Sie das Bild unter CC BY 2.0 hochgeladen haben! Thank you very much!
-
↑Bild VB23006.JPG:
File:Amphiura constricta 02.jpg und hier von Julian Finn, Museums Victoria
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
-
Michael Eitel,
Hans-Jürgen Osigus, Rob DeSalle, Bernd Schierwater: Global Diversity of the Placozoa. In: PLoS One, Published: April 2, 2013, https://doi.org/10.1371/journal.pone.0057131 ( Volltext)
- 11.1 ↑Bild VB23003.PNG:
Ausschnitt aus: File:Trichoplax adhaerens photograph.png und hier von Bernd Schierwater - aus Michael Eitel, Hans-Jürgen Osigus, Rob DeSalle, Bernd Schierwater: Global Diversity of the Placozoa. In: PLoS One, Published: April 2, 2013, https://doi.org/10.1371/journal.pone.0057131 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 11.1 ↑Bild VB23003.PNG:
Ausschnitt aus:
-
Yoshinori Shichida,
Take Matsuyama: Evolution of opsins and phototransduction. In: Philosophical Transactions of the Royal Society of London, (2009) 364, 2881–2895, PMID: 19720651, doi:10.1098/rstb.2009.0051 ( Volltext)
- 12.1 ↑Bild O00071901.GIF:
"Stammbaum der Opsine" von Kersti Nebelsiek
Informationen nach: Yoshinori Shichida, Take Matsuyama: Evolution of opsins and phototransduction. In: Philosophical Transactions of the Royal Society of London, (2009) 364, 2881–2895, PMID: 19720651, doi:10.1098/rstb.2009.0051 ( Volltext)
Lizenz: GNU 1.2, CC BY-SA 4.0, CC BY-SA 3.0, CC BY-SA 2.5, CC BY-SA 2.0, CC BY-SA 1.0
- 12.1 ↑Bild O00071901.GIF:
"Stammbaum der Opsine" von Kersti Nebelsiek
-
Zuzana Musilova,
Fabio Cortesi, Michael Matschiner, Wayne I. L. Davies, Jagdish Suresh Patel, Sara M. Stieb, Fanny de Busserolles, Martin Malmstrøm, Ole K. Tørresen, Celeste J. Brown, Jessica K. Mountford, Reinhold Hanel, Deborah L. Stenkamp, Kjetill S. Jakobsen, Karen L. Carleton, Sissel Jentoft, Justin Marshall, Walter Salzburger: Vision using multiple distinct rod opsins in deep-sea fishes. In: Science, 2019 May 10; 364(6440): 588–592, doi: 10.1126/science.aav4632, PMID: 31073066 ( Volltext)
-
Rui Borges,
Warren E. Johnson, Stephen J. O’Brien, Cidália Gomes, Christopher P. Heesy, Agostinho Antunes: Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. In: BMC Genomics, volume 19, Article number: 121 (2018) ( Volltext)
- 14.1 ↑Bild O000719.GIF:
Fig. 3 aus Rui Borges, Warren E. Johnson, Stephen J. O’Brien, Cidália Gomes, Christopher P. Heesy, Agostinho Antunes: Adaptive genomic evolution of opsins reveals that early mammals flourished in nocturnal environments. In: BMC Genomics, volume 19, Article number: 121 (2018) ( Volltext)
Mitverwendet wurden folgende Bilder File:Shennongjia virgin forest.jpg von User:Evilbish von Wikimedia Commons
und File:Pristeroognathus DB.jpg von Участник:ДиБгд aus der russischen Wikipedia
Vielen Dank, daß Sie das Bild unter CC BY-SA 3.0 hochgeladen haben! Thank you very much!
- 14.1 ↑Bild O000719.GIF:
-
Nicole Y. Leung,
Craig Montell: Unconventional roles of opsins. In: Annual review of cell and developmental biology, Author manuscript; available in PMC 2018 Oct 6, Published in final edited form as: Annual review of cell and developmental biology, 2017 Oct 6; 33: 241–264, doi: 10.1146/annurev-cellbio-100616-060432, PMID: 28598695 ( Volltext)
-
↑Bild VB23004.PNG:
File:Chlamydomonas reinhardtii vector scheme.svg von User:Nefronus von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC0 1.0 hochgeladen hast! Thank you very much!
-
Daria N. Shalaeva,
Michael Y. Galperin, Armen Y. Mulkidjanian: Eukaryotic G protein-coupled receptors as descendants of prokaryotic sodium-translocating rhodopsins. In: Biology direct, volume 10, Article number: 63 (2015) ( Volltext)
- 17.1 ↑Bild VB230.GIF:
Fig. 2 aus: Daria N. Shalaeva,
Michael Y. Galperin, Armen Y. Mulkidjanian: Eukaryotic G protein-coupled receptors as descendants of prokaryotic sodium-translocating rhodopsins. In: Biology direct, volume 10, Article number: 63 (2015) ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 17.1 ↑Bild VB230.GIF:
-
Mansi Srivastava,
Emina Begovic, Jarrod Chapman, Nicholas H. Putnam, Uffe Hellsten, Takeshi Kawashima, Alan Kuo, Therese Mitros, Asaf Salamov, Meredith L. Carpenter, Ana Y. Signorovitch, Maria A. Moreno, Kai Kamm, Jane Grimwood, Jeremy Schmutz, Harris Shapiro, Igor V. Grigoriev, Leo W. Buss, Bernd Schierwater, Stephen L. Dellaporta, Daniel S. Rokhsar: The Trichoplax genome and the nature of placozoans. In: Nature, volume 454, pages955–960 (2008) ( Volltext)
-
Jörg Peter Ewert,
Sabine Beate Ewert:
B69.11
Wahrnehmung. (1981) Heidelberg: Quelle und Meyer, ISBN 3-494-01060-9
-
Hans-Eckhard Gruner (Hrsg):
B167.12
Lehrbuch der Speziellen Zoologie, Band I Wirbellose Tiere, 2.Teil: Cnidaria, Ctenophora, Mesozoa, Plathelminthes, Nemertini, Entoprocta, Nemathelminthes, Priapulida (1993) Stuttgart, New York: Gustav Fischer Verlag, ISBN: 3334604748
-
Roberto Feuda,
Sinead C. Hamilton, James O. McInerney, Davide Pisani: Metazoan opsin evolution reveals a simple route to animal vision. In: PNAS, November 13, 2012 109 (46) 18868-18872; https://doi.org/10.1073/pnas.1204609109 ( Volltext)
-
Ajna S. Rivera,
Nuri Ozturk, Bryony Fahey, David C. Plachetzki, Bernard M. Degnan, Aziz Sancar, Todd H. Oakley: Blue-light-receptive cryptochrome is expressed in a sponge eye lacking neurons and opsin. In: The Journal of Experimental Biology, 2012 215: 1278-1286; doi: 10.1242/jeb.067140 ( Volltext)
-
Maja Adamska,
Sandie M. Degnan, Kathryn M. Green, Marcin Adamski, Alina Craigie, Claire Larroux, Bernard M. Degnan: Wnt and TGF-β Expression in the Sponge Amphimedon queenslandica and the Origin of Metazoan Embryonic Patterning. In: PLoS One, 2007; 2(10): e1031, PMID: 17925879 ( Volltext)
- 23.1 ↑Bild VB23007.JPG:
Ausschnitte aus: Figure 1. Amphimedon queenslandica life cycle and embryonic development. In: Maja Adamska, Sandie M. Degnan, Kathryn M. Green, Marcin Adamski, Alina Craigie, Claire Larroux, Bernard M. Degnan: Wnt and TGF-β Expression in the Sponge Amphimedon queenslandica and the Origin of Metazoan Embryonic Patterning. In: PLoS One, 2007; 2(10): e1031, PMID: 17925879 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 23.2 ↑Bild VB23008.JPG:
Ausschnitt aus: Figure 1. Amphimedon queenslandica life cycle and embryonic development. Aus: Maja Adamska, Sandie M. Degnan, Kathryn M. Green, Marcin Adamski, Alina Craigie, Claire Larroux, Bernard M. Degnan: Wnt and TGF-β Expression in the Sponge Amphimedon queenslandica and the Origin of Metazoan Embryonic Patterning. In: PLoS One, 2007; 2(10): e1031, PMID: 17925879 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 23.1 ↑Bild VB23007.JPG:
Ausschnitte aus: Figure 1. Amphimedon queenslandica life cycle and embryonic development. In:
-
Hiroshi Suga,
Volker Schmid, Walter J. Gehring: Evolution and Functional Diversity of Jellyfish Opsins. In: Current biology, Volume 18, Issue 1, 8 January 2008, Pages 51-55 https://doi.org/10.1016/j.cub.2007.11.059 ( Volltext)
-
Natasha Picciani,
Jamie R. Kerlin, Noemie Sierra, Andrew J. M. Swafford, M. Desmond Ramirez, Nickellaus G. Roberts, Johanna T. Cannon, Marymegan Daly, Todd H. Oakley: Prolific Origination of Eyes in Cnidaria with Co-option of Non-visual Opsins. In: Current biology, Volume 28, 2413–2419, August 6, 2018 ( Volltext)
-
David C. Plachetzki,
Bernard M. Degnan, Todd H. Oakley: The Origins of Novel Protein Interactions during Animal Opsin Evolution. In: PLoS One, 2(10): e1054, Published: October 17, 2007. https://doi.org/10.1371/journal.pone.0001054 ( Volltext)
-
Ouria Dkhissi-Benyahya,
Camille Rieux, Roelof A. Hut, Howard M. Cooper: Immunohistochemical Evidence of a Melanopsin Cone in Human Retina. In: Investigative Ophthalmology & Visual Science, April 2006, Vol. 47, No. 4, 1636-1641, PMID: 16565403, DOI: 10.1167/iovs.05-1459 ( Volltext)
-
Annette E. Allen,
Franck P. Martial, Robert J. Lucas: Form vision from melanopsin in humans. In: Nature Communications, volume 10, Article number: 2274 (2019) ( Volltext)
-
Kirk Haltaufderhyde,
Rana N. Ozdeslik, Nadine L. Wicks, Julia A. Najera, Elena Oancea: Opsin Expression in Human Epidermal Skin. In: Photochemistry and Photobiology, 2015, 91: 117 – 123 PMID: 25267311, DOI: 10.1111/php.12354 ( Volltext 1, 2)
-
Moe Tsutsumi,
Kazuyuki Ikeyama, Sumiko Denda, Jotaro Nakanishi, Shigeyoshi Fuziwara, Hirofumi Aoki, Mitsuhiro Denda: Expressions of rod and cone photoreceptor-like proteins in human epidermis. In: Experimental Dermatology, 18, (2009) 567–570, PMID: 19493002, DOI: 10.1111/j.1600-0625.2009.00851.x ( Volltext)
-
↑Bild VB23009.JPG:
File:Diretmus argenteus1.jpg, oder Pl V von Emma Kissling, gestorben 1913, aus Albert I, Prinz von Monaco, Jules de Guerne, Jules Richard: B167.13 Résultats des campagnes scientifiques accomplies sur son yacht par Albert Ier, prince souverain de Monaco. Fascicule XXXV Poissons provenant des campagnes du yacht Princesse-Alice (1901-1910) (1911) Imprimierie de Monaco
Das Bild ist gemeinfrei, da die urheberrechtliche Schutzfrist abgelaufen ist
-
↑Bild O00071905.PNG:
File:1L9H (Bovine Rhodopsin) 2.png von Sven Jähnichen (User:S. Jähnichen von Wikimedia Commons)
Vielen Dank, daß Du das Bild als gemeinfrei veröffentlicht hast, so weit das rechtlich möglich ist! Thank you very much!
-
Krzysztof Palczewski,
Takashi Kumasaka, Tetsuya Hori, Craig A. Behnke, Hiroyuki Motoshima, Brian A. Fox, Isolde Le Trong, David C. Teller, Tetsuji Okada, Ronald E. Stenkamp, Masaki Yamamoto, Masashi Miyano: Crystal Structure of Rhodopsin: A G Protein–Coupled Receptor. In: Science, 04 Aug 2000: Vol. 289, Issue 5480, pp. 739-745, DOI: 10.1126/science.289.5480.739 ( Volltext)
-
↑Bild O00071907.PNG:
File:Phototransduction.png von Jason J. Corneveaux (User:Caddymob von Wikimedia Commons)
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY 3.0 hochgeladen hast! Thank you very much!
-
Joseph T. Ortega,
Beata Jastrzebska: The Retinoid and Non-Retinoid Ligands of the Rod Visual G Protein-Coupled Receptor. In: International Journal of Molecular Sciences 2019, 20, 6218; doi:10.3390/ijms20246218 ( Volltext 1, 2)
- 35.1 ↑Bild O00071910.PNG:
Abwandung von Figure 1. aus Joseph T. Ortega,
Beata Jastrzebska: The Retinoid and Non-Retinoid Ligands of the Rod Visual G Protein-Coupled Receptor. In: International Journal of Molecular Sciences 2019, 20, 6218; doi:10.3390/ijms20246218 ( Volltext 1, 2), umgedreht, neu beschriftet und Membran eingezeichnet durch Kersti Nebelsiek
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 35.1 ↑Bild O00071910.PNG:
Abwandung von Figure 1. aus
-
Fanny de Busserolles,
Lily Fogg, Fabio Cortesi, Justin Marshall: The exceptional diversity of visual adaptations in deep-sea teleost fishes. In: Seminars in Cell & Developmental Biology, 11 Jun 2020, 106:20-30, DOI: 10.1016/j.semcdb.2020.05.027, PMID: 32536437 ( Volltext)
-
Ronald H. Douglas,
Martin J. Genner, Alan G. Hudson, Julian C. Partridge, Hans-Joachim Wagner: Localisation and origin of the bacteriochlorophyll-derived photosensitizer in the retina of the deep-sea dragon fish Malacosteus niger. In: Scientific Reports, volume 6, Article number: 39395 (2016) ( Volltext)
- 37.1 ↑Bild VB23005.PNG:
Figure 1 aus Ronald H. Douglas, Martin J. Genner, Alan G. Hudson, Julian C. Partridge, Hans-Joachim Wagner: Localisation and origin of the bacteriochlorophyll-derived photosensitizer in the retina of the deep-sea dragon fish Malacosteus niger. In: Scientific Reports, volume 6, Article number: 39395 (2016) ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 37.1 ↑Bild VB23005.PNG:
Figure 1 aus
-
Peter J. Herring,
Celia Cope: Red bioluminescence in fishes: on the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. In: Marine Biology, volume 148, pages383–394(2005) ( Volltext)
- 38.1 ↑Bild VB23013.JPG:
File:Malacosteus niger cam.jpg aus: Peter J. Herring, Celia Cope: Red bioluminescence in fishes: on the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. In: Marine Biology, volume 148, pages383–394(2005) ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY-SA 3.0 hochgeladen haben! Thank you very much!
- 38.1 ↑Bild VB23013.JPG:
-
Katarina Ondrusova,
Mohammad Fatehi, Amy Barr, Zofia Czarnecka, Wentong Long, Kunimasa Suzuki, Scott Campbell, Koenraad Philippaert, Matthew Hubert, Edward Tredget, Peter Kwan, Nicolas Touret, Martin Wabitsch, Kevin Y. Lee, Peter E. Light: Subcutaneous white adipocytes express a light sensitive signaling pathway mediated via a melanopsin/TRPC channel axis. In: Scientific Reports, 7, Article number: 16332 (2017) , DOI:10.1038/s41598-017-16689-4 ( Volltext)
-
Junya Kusumoto,
Makoto Takeo, Kazunobu Hashikawa, Takahide Komori, Takashi Tsuji, Hiroto Terashi, Shunsuke Sakakibara: OPN4 belongs to the photosensitive system of the human skin. In: Genes to cells, Volume25, Issue3, March 2020, Pages 215-225, PMID: 31989708, DOI: 10.1111/gtc.12751 ( Volltext)
-
Gautam Sikka,
G. Patrick Hussmann, Deepesh Pandey, Suyi Cao, Daijiro Hori, Jong Taek Park, Jochen Steppan, Jae Hyung Kim, Viachaslau Barodka, Allen C. Myers, Lakshmi Santhanam, Daniel Nyhan, Marc K. Halushka, Raymond C. Koehler, Solomon H. Snyder, Larissa A. Shimoda, Dan E. Berkowitza: Melanopsin mediates light-dependent relaxation in blood vessels. In: PNAS, December 16, 2014 111 (50) 17977-17982; first published November 17, 2014; doi: 10.1073/pnas.1420258111 ( Volltext 1, 2)
-
Bhanu Iyengar:
The melanocyte photosensory system in the human skin. In: SpringerPlus, volume 2, Article number: 158 (2013) ( Volltext)
-
Susie Suh,
Elliot H. Choi, Natasha Atanaskova Mesinkovska: The expression of opsins in the human skin and its implications for photobiomodulation: A Systematic Review. In: Photodermatology, photoimmunology & photomedicine, Volume36, Issue5, September 2020, Pages 329-338 ( Volltext)
-
↑Bild VB23011.JPG:
Beschriftete und verkleinerte Version von: File:Retina -- high mag.jpg von User:Librepath von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23012.JPG:
File:Malacosteus niger cam.jpg oder Pl. III von Emma Kissling, gestorben 1913, aus Albert I, Prinz von Monaco, Jules de Guerne, Jules Richard: B167.13 Résultats des campagnes scientifiques accomplies sur son yacht par Albert Ier, prince souverain de Monaco. Fascicule XXXV Poissons provenant des campagnes du yacht Princesse-Alice (1901-1910) (1911) Imprimierie de Monaco
Das Bild ist gemeinfrei, da die urheberrechtliche Schutzfrist abgelaufen ist
-
↑Bild VB23010.JPG:
File:Beroe abyssicola 04.jpg von User:Σ64 von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY 3.0 hochgeladen hast! Thank you very much!
-
Ronald H. Douglas,
Conrad W. Mullineaux, Julian C. Partridge: Long-wave sensitivity in deep-sea stomiid dragonfish with far-red bioluminescence: evidence for a dietary origin of the chlorophyll-derived retinal photosensitizer of Malacosteus niger. In: Philosophical Transactions of the Royal Society of London, B (2000) 355, 1269-1272, doi 10.1098/rstb.2000.0681 ( Volltext)
-
↑Bild VA08301.PNG:
File:Spectre.svg von User:Tatoute und User:Phrood von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0, CC BY-SA 2.5, CC BY-SA 2.0, CC BY-SA 1.0 hochgeladen hast! Thank you very much!
-
Morgane Bonadè,
Atsushi Ogura, Erwan Corre, Yann Bassaglia, Laure Bonnaud-Ponticelli: Diversity of Light Sensing Molecules and Their Expression During the Embryogenesis of the Cuttlefish (Sepia officinalis). In: Frontiers in Physiology, 7:660, 29 September 2020 DOI: 10.3389/fphys.2020.521989 ( Volltext)
-
Justine J. Allen,
Lydia M. Mäthger, Kendra C. Buresch, Thomas Fetchko, Meg Gardner, Roger T. Hanlon: Night vision by cuttlefish enables changeable camouflage. In: The Journal of Experimental Biology, Volume 213, Issue 23, December 2010 ( Volltext)
-
Martin H. Moynihan,
Arcadio F. Rodaniche: Chapter 14. Communication, crypsis, and mimicry among cephalopods. ( Volltext) In: Thomas A. Sebeok: B141.21 How Animals Communicate. (1977) Bloomington, Indiana USA: Indiana University Press, ISBN 978-0-253-05093-9 ( Volltext)
-
↑Bild VB23014.JPG:
File:2018.04.07.-03-Kirschgartshaeuser Schlaege Mannheim--Gemeiner Furchenschwimmer-Weibchen.jpg von Andreas Eichler ( User:Hockei von Wikimedia Commons)
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23006.PNG:
File:Origin of Vertebrates Fig 035.png oder hier aus: Walter Holbrook Gaskell: B86.12 The origin of vertebrates. (1908) London, New York, Bombay, Calcutta, Longmans, Green, and co. ( Volltext)
Das Bild ist aufgrund seines Alters gemeinfrei
-
↑Bild VB23015.JPG:
File::Acilius.sulcatus5.-.lindsey.jpg von James K. Lindsey The Ecology of Commanster
Vielen Dank, daß Du das Bild unter CC BY-SA 2.5, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23016.JPG:
File:Washington DC Zoo - Sepia officinalis 2.jpg von Jarek Tuszyński ( User:Jarekt von Wikimedia Commons)
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23017.JPG:
File:Herklots 1859 I 1 Sepia officinalis - dier.jpg aus: Janus Adrian Herklots B167.14 De Weekdieren van Nederland. Natuurlijke Historie van Nederland. De Dieren van Nederland, Weekdieren. (1859) Haarlem: Kruseman
Das Bild ist aufgrund seines Alters gemeinfrei.
-
↑Bild VB23018.JPG:
File:Sepia officinalis 2648.jpg von User:Amada44 von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23019.JPG:
File:Sepia officinalis 2651.jpg von User:Amada44 von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23020.JPG:
File:Sepia officinalis.001 - Aquarium Finisterrae.jpg von Fernando Losada Rodríguez ( User:Drow male von Wikimedia Commons)
Vielen Dank, daß Du das Bild unter CC BY-SA 4.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23021.JPG:
File:Gewöhnlicher Tintenfisch.jpg von Marco Almbauer ( User:A,Ocram von Wikimedia Commons)
Vielen Dank, daß Du das Bild unter CC0 1.0 hochgeladen hast! Thank you very much!
-
↑Bild VB23022.JPG:
File:Sepia officinalis - Carantec-2.jpg von Matthieu Sontag ( User:Mirgolth) von Wikimedia Commons
Vielen Dank, daß Du das Bild unter GNU 1.2, CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Wen-Sung Chung,
N. Justin Marshall: Complex Visual Adaptations in Squid for Specific Tasks in Different Environments. In: Frontiers in Physiology, 8:105. 2017 doi: 10.3389/fphys.2017.00105 ( Volltext)
-
Anne-Sophie Darmaillacq,
Nawel Mezrai, Caitlin E. O'Brien, Ludovic Dickel: Visual Ecology and the Development of Visually Guided Behavior in the Cuttlefish. In: Frontiers in Physiology, 8:402, 13 June 2017, doi: 10.3389/fphys.2017.00402 ( Volltext)
- 63.1 ↑Bild VB23023.JPG:
Figure 5 aus Anne-Sophie Darmaillacq, Nawel Mezrai, Caitlin E. O'Brien, Ludovic Dickel: Visual Ecology and the Development of Visually Guided Behavior in the Cuttlefish. In: Frontiers in Physiology, 8:402, 13 June 2017, doi: 10.3389/fphys.2017.00402 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
- 63.1 ↑Bild VB23023.JPG:
Figure 5 aus
-
Keri V. Langridge:
Symmetrical crypsis and asymmetrical signalling in the cuttlefish Sepia officinalis. In: Proceedings of the Royal Society of London Series B. (2006) 273, 959–967 doi:10.1098/rspb.2005.3395 ( Volltext)
-
Deanna Panetta,
Kendra C. Buresch, Roger T. Hanlon: Dynamic masquerade with morphing three-dimensional skin in cuttlefish. In: Biology Letters 29 March 2017, Volume 13 Issue 3 PMID: 28356412, DOI: 10.1098/rsbl.2017.0070 ( Volltext)
-
Chuan-Chin Chiao,
J. Kenneth Wickiser, Justine J. Allen, Brock Genter, Roger T. Hanlon: Hyperspectral imaging of cuttlefish camouflage indicates good color match in the eyes of fish predators. In: PNAS, 2011 May 31;108(22):9148-53. Epub 2011 May 16. PMID: 21576487, DOI: 10.1073/pnas.1019090108 ( Volltext)
-
↑Bild VB23024.JPG:
Figure 1. aus Boudjema Imarazene, Aude Andouche, Yann Bassaglia, Pascal-Jean Lopez, Laure Bonnaud-Ponticelli: Eye Development in Sepia officinalis Embryo: What the Uncommon Gene Expression Profiles Tell Us about Eye Evolution. In: Frontiers in Physiology, 8:613, 24 August 2017, doi: 10.3389/fphys.2017.00613 ( Volltext)
Vielen Dank, daß Sie das Bild unter CC BY 4.0 hochgeladen haben! Thank you very much!
-
Lydia M. Mäthger,
Alexandra Barbosa, Simon Miner, Roger T. Hanlon: Color blindness and contrast perception in cuttlefish (Sepia officinalis) determined by a visual sensorimotor assay. In: Vision Research, Volume 46, Issue 11, May 2006, Pages 1746-1753 doi:10.1016/j.visres.2005.09.035 ( Volltext)
-
Sarah Zylinski,
Daniel Osorio, Adam J. Shohet: Cuttlefish camouflage: context-dependent body pattern use during motion. In: Proceedings of the Royal Society of London Series B. 276, 3963–3969 doi:10.1098/rspb.2009.1083, PMID: 19692411 ( Volltext)
-
David Lagman,
Daniel Ocampo Daza, Jenny Widmark, Xesús M. Abalo, Görel Sundström, Dan Larhammar: The vertebrate ancestral repertoire of visual opsins, transducin alpha subunits and oxytocin/vasopressin receptors was established by duplication of their shared genomic region in the two rounds of early vertebrate genome duplications. In: BMC Evolutionary Biology, volume 13, Article number: 238 (2013) ( Volltext)
-
Marcus Robert Jones,
Sten Grillner, Brita Robertson: Selective projection patterns from subtypes of retinal ganglion cells to tectum and pretectum: distribution and relation to behavior. In: Journal of Comparative Neurology 517:257–275 (2009) DOI: 10.1002/cne.22203, PMID: 19760658 ( Volltext)
-
Gordon L. Fain:
Lamprey vision: Photoreceptors and organization of the retina. In: Seminars in Cell & Developmental Biology, Volume 106, October 2020, Pages 5-11 https://doi.org/10.1016/j.semcdb.2019.10.008 ( Volltext)
-
Jennifer M. Enright,
Matthew B. Toomey, Shin-ya Sato, Shelby E. Temple, James R. Allen, Rina Fujiwara, Valerie M. Kramlinger, Leslie D. Nagy, Kevin M. Johnson, Yi Xiao, Martin J. How, Stephen L. Johnson, Nicholas W. Roberts, Vladimir J. Kefalov, F. Peter Guengerich, Joseph C. Corbo: Cyp27c1 Red-Shifts the Spectral Sensitivity of Photoreceptors by Converting Vitamin A1 into A2. In: Current biology, Volume 25, ISSUE 23, P3048-3057, December 07, 2015 ( doi: 10.1016/j.cub.2015.10.018)
-
Omar B. Sanchez-Reyes,
Aidan L. G. Cooke, Dale B. Tranter, Dawood Rashid, Markus Eilers, Philip J. Reeves, Steven O. Smith: G Protein-Coupled Receptors Contain Two Conserved Packing Clusters. In: Biophysical Journal, 2017 Jun 6; 112(11): 2315–2326. DOI: 10.1016/j.bpj.2017.04.051 ( Volltext)
-
Xavier Deupi,
Patricia Edwards, Ankita Singhal, Benjamin Nickle, Daniel Oprian, Gebhard F. X. Schertler, Jörg Standfuss: Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. In: PNAS, January 3, 2012; vol. 109, no. 1, 119–124 ( Volltext)
-
Sarah Zylinski,
Daniel Osorio, Sönke Johnsen: Cuttlefish see shape from shading, finetuning coloration in response to pictorial depth cues and directional illumination. In: Proceedings of the Royal Society of London, B (2016) 283: 20160062. ( Volltext)
-
Lydia M. Mäthger,
Steven B. Roberts, Roger T. Hanlon: Evidence for distributed light sensing in the skin of cuttlefish, Sepia officinalis. In: Biology Letters (2010) 6, 600–603, doi:10.1098/rsbl.2010.0223 ( Volltext 1, 2)
-
Yi-Hsin Lee,
Hong Young Yan, Chuan-Chin Chiao: Effects of early visual experience on the background preference in juvenile cuttlefish Sepia pharaonis. In: Biology Letters (2012) 8, 740–743, doi:10.1098/rsbl.2012.0398 ( Volltext)
-
Kendra C. Buresch,
Kimberly M. Ulmer, Derya Akkaynak, Justine J. Allen, Lydia M. Mäthger, Mario Nakamura, Roger T. Hanlon: Cuttlefish adjust body pattern intensity with respect to substrate intensity to aid camouflage, but do not camouflage in extremely low light. In: Journal of Experimental Marine Biology and Ecology, Volume 462, January 2015, Pages 121-126 ( Volltext)
-
Alexander L. Stubbs,
Christopher W. Stubbs: Spectral discrimination in color blind animals via chromatic aberration and pupil shape. In: PNAS, July 19, 2016, vol. 113, no. 29, 8206–8211, https://doi.org/10.1073/pnas.1524578113 ( Volltext)
-
↑Bild O00071913.PNG:
File:Lens6a.svg von User:Pitel und User:DrBob von Wikimedia Commons
Vielen Dank, daß Du das Bild unter CC BY-SA 3.0 hochgeladen hast! Thank you very much!
-
Dongmin Keum,
Kyung-Won Jang, Daniel S. Jeon, Charles S. H. Hwang, Elke K. Buschbeck, Min H. Kim, Ki-Hun Jeong: Xenos peckii vision inspires an ultrathin digital camera. In: Light: Science & Applications, volume 7, Article number: 80 (2018) ( Volltext)
- 82.1
↑Bild VB23008.PNG:
Fig. 1: Natural Xenos peckii eye and the biological inspiration for the ultrathin digital camera ( File:Xenos peckii eye and ultrathin digital camera..png) Dongmin Keum, Kyung-Won Jang, Daniel S. Jeon, Charles S. H. Hwang, Elke K. Buschbeck, Min H. Kim, Ki-Hun Jeong: Xenos peckii vision inspires an ultrathin digital camera. In: Light: Science & Applications, volume 7, Article number: 80 (2018) ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY 4.0 hochgeladen hast! Thank you very much!
- 82.2
↑Bild VB23009.PNG:
Fig. 1: Natural Xenos peckii eye and the biological inspiration for the ultrathin digital camera ( File:Xenos peckii eye and ultrathin digital camera..png) Dongmin Keum, Kyung-Won Jang, Daniel S. Jeon, Charles S. H. Hwang, Elke K. Buschbeck, Min H. Kim, Ki-Hun Jeong: Xenos peckii vision inspires an ultrathin digital camera. In: Light: Science & Applications, volume 7, Article number: 80 (2018) ( Volltext)
Vielen Dank, daß Du das Bild unter CC BY 4.0 hochgeladen hast! Thank you very much!
- 82.1
↑Bild VB23008.PNG:
-
↑Bild VB23025.JPG:
File:Purple sea urchin (42974674532).jpg (oder hier) von Flickr-User brewbooks
CC BY 2.0 Vielen Dank! Thank you very much!
-
Yale J. Passamaneck,
Nina Furchheim, Andreas Hejnol, Mark Q. Martindale, Carsten Lüter: Ciliary photoreceptors in the cerebral eyes of a protostome larva. BZ467. EvoDevo, volume 2, Article number: 6 (2011) ( Volltext)
- 84.1 ↑Bild VB23027.JPG:
Figure 1 in Yale J. Passamaneck,
Nina Furchheim, Andreas Hejnol, Mark Q. Martindale, Carsten Lüter: Ciliary photoreceptors in the cerebral eyes of a protostome larva. BZ467. EvoDevo, volume 2, Article number: 6 (2011) ( Volltext)
CC BY 2.0 Vielen Dank! Thank you very much!
- 84.1 ↑Bild VB23027.JPG:
Figure 1 in
-
M. Desmond Ramirez,
Autum N. Pairett, M. Sabrina Pankey, Jeanne Marie Serb, Daniel I. Speiser, Andrew J. M. Swafford, Todd H. Oakley: The Last Common Ancestor of Most Bilaterian Animals Possessed at Least Nine Opsins. In: Genome Biology and Evolution, Volume 8, Issue 12, December 2016, Pages 3640–3652, ( Volltext)
-
Thomas W. Cronin,
Sönke Johnsen: Extraocular, Non-Visual, and Simple Photoreceptors: An Introduction to the Symposium. In: Integrative and Comparative Biology, volume 56, number 5, pp. 758–763, doi:10.1093/icb/icw106 ( Volltext)
-
Karin Nordström,
Rita Wallén, Jamie Seymour, Dan Nilsson: A simple visual system without neurons in jellyfish larvae. In: Proceedings of the Royal Society of London B (2003) 270, 2349–2354, DOI 10.1098/rspb.2003.2504 ( Volltext)
-
Jorge Alves Audino,
Jeanne Marie Serb, José Eduardo Amoroso Rodriguez Marian: Hard to get, easy to lose: Evolution of mantle photoreceptor organs in bivalves (Bivalvia, Pteriomorphia). In: Evolution (2020) 74-9: 2105–2120, PMID: 32716056, DOI: 10.1111/evo.14050 ( Volltext)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_999_(30695685804).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_2.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}